Навигация
Вход в систему

Nothobranchius microlepis (Vinciguerra, 1897)

Первое описание
Vinciguerra, D. - 1897 - "Pesci raccolti dal la V. Bottego, durante la sua seconda spedizione nelle regioni dei Somalia e dei Galla." - Annali del Museo Civice di Storia Naturale di Genoa 2,17: 343-364.
Оригинальное описание этого вида утверждает, что N.microlepis, впервые выловленный V. Bottego, отличается от остальных известных видов рода следующими чертами : (1) большим количеством мелких чешуек на теле, (2) наличием тёмной вертикальной полоски, идущей сверху головы через глаз до угла рта, (3) большим межглазничным расстоянием, (4) заметно выпуклой линией спины и (5) сдвинутым дальше назад спинным плавником.
Типовое место обитания
"Stagno Saha, sotto il monte Egberta, tra Brava e Lugh" = стандартный пруд у подножия горы Egberta (ныне Egherta) между Brava и Lugh (ныне Lugh Ganana) в южной Сомали. Горы Egherta Mountains северозападнее Brava питает водой пересыхающая река, известная под названием Madagoi. Особи этого вида были впервые выловлены во временных водоёмах в системе этой реки.
Дополнительные места вылова этого вида находятся юго-восточной Сомали, включая Warfa, Durbane, Bur Eibi/ Kaisany, Hogay, Egherta, и Dekta.
Места поимки в северо-восточной Кении включают среднюю часть дренажной системы Tana, а так же вдоль дороги из Garsen в Garissa (Willert, 1984).
Синонимы
- Fundulus microlepis Vinciguerra, (1897): 356
Fundulopanchax microlepis Meinken (1930): 117-119, 597-598
Nothobranchius microlepis Myers (1933): 180,185
Принимая во внимание различия между этим видом и "стандартными" Nothobranchius в форме тела взрослых самцов (горбатые), биологии (сильно отличающееся число хромосом по данным Scheel, чёрная вертикальная полоса, идущая через глаз, большое количество мелких беспорядочно расположенных чешуй, окраска) и поведению (биологич размножения), может так случиться, что в будущем для этого вида будет образован самостоятельный подрод или отдельный род.
Родственные связи
Wildekamp & Haas (1992) отмечают, что характеристики (большое количество мелких чешуй, тёмная полоса на глазу, большое межглазничное расстояние, горб у самца и далеко расположенный спинной плавник), использованные Vinciguerra выделить N.microlepis из остальных видов рода являются синапоморфными для этого вида и N.fasciatus и N.bojiensis. Дополнительные характеристики: выступ верхней челюсти напротив зрачка, длинные и близко посаженные жаберные тычинки и поведение при ухаживании. Так как эти характеристики неизвестны неизвестны среди Aplocheilidae, Wildekamp & Haas (1992) сочли их синапоморфными для этой группы видов.
По Wildekamp & Haas (1992) некоторые из этих синапоморфных характеристик (большое количество беспорядочно расположенных чешуек, большое межглазничное расстояние, горб у самца, далеко расположенный спинной плавник и удлинённая верхняя челюсть на уровне зрачка) так же проявляется у P.ocellatus. Но последний отличается от N.microlepis, N.fasciatus и N.bojiensis наличием меньшего количества жаберных тычинок (11 - 12) на первой жаберной дуге, что не отличается от N.jubbi и других Nothobranchius, а так же поведение во время ухаживания не отличается от других видов Nothobranchius.
Seegers (1985) описал монотипичный род (= включающий только один вид) Paranothobranchius (типовой вид: N.ocellatus Seegers, 1985) основываясь на экземпляре этого вида и сделанном с него слайде. В этом неполном описании упоминается более крупное рыло с точеками, более плоская и удлинённая голова, более удлинённое и сравнительно тонкое тело, большее расстояние до брюшных плавников и расположенные ближе к заднему концу тела спинной и анальнвй плавники, а так же большое количество мелких чешуй в продольном ряду, как характеристики, отличающие этот род от родственного ему рода Nothobranchius. Но не было приведено сравнения с характеристиками, уникальными для Nothobranchius Parenti (1981): интераркуальный хрящ (interarcual cartilage) прикреплён непосредственно к хрящу верхнему отделу второй жаберной дуги, овальная (а не округлая) икра и не закрытый преоперкулярный канал органов боковой линии. Из этих характеристик все повторяются у Paranothobranchius. Wildekamp & Haas (1992) заметили, что как овальные, так и округлые икринки могут быть обнаружены у всех видов Nothobranchius, что уменьшает ценность этой определительной характеристики. К тому же они нашли у всех видов рода Nothobranchius, включая P.ocellatus, равномерно расположенные, крючкообразной формы короткие филаменты на внешней оболочке икринки. Эта характеристика уникальна среди Aplocheilidae и может использоваться как определительная для рода Nothobranchius.
Дополнительные характеристики, приводимые Seegers (1985) для выделения P.ocellatus из рода Nothobranchius следующие: менее развитые ктеноидные шипики, более тонкая эпидермальная ткань, покрывающая верхние чешуи на голове, менее развитая супраорбитальная система органов боковой линии, выступающая перед передним краем зрачка верхняя челюстьотсутствие недостатка кожи, покрывающей заднюю часть нижней челюсти, очень заметный переход с изгибом от головы к спине, отсутствие яркой окраски у самцов и наличие глазчатого пятна на хвосте (=тёмное округлое пятно). N.microlepis, N.fasciatus и N.bojiensis соответствуют этим характеристикам. Wildekamp & Haas (1992) обнаружили, что непокрытая задняя часть нижней челюсти присутствует и у исследованных P.ocellatus из Bagamoyo, разведённых в аквариуме, а так же что отсутствие яркой окраски у самцов - характерная черта для ювенильных особей.
Таким образом Wildekamp & Haas (1992) предположили, что N.microlepis, N.bojiensis и N.ocellatus являются представителями монофилетичной линии, предворительно определённой как подрод Paranothobranchius внутри рода Nothobranchius. Он определяется следующими общими характерными (синапоморфными) признаками: мелкие, беспорядочно расположенные чешуи, большой рот с большим количеством зубов, длинная голова, горб у самца и дальше к хвосту расположенные спинной и анальный плавники. В этой линии Wildekamp & Haas (1992) сочли N.ocellatus наиболее примитивным представителем, так как он проявляет и плезиоморфные для остальных Nothobranchius признаки: наличие чёткого полового диморфизма в окраске, отсутствие тёмной вертикальной полосы через глаз, низкое количество жаберных тычинок на первой жаберной дуге и поведение во время ухаживания самца за самкой. Нзкое количество жаберных тычинок указывает на другое кормовое предпочтение N.ocellatus, который является хищником, питающимся мелкой рыбой (например другими видами Nothobranchius). Wildekamp & Haas (1992) далее предположили неясными межродственные связи между Paranothobranchius и Nothobranchius, так как они не могли выделить набор признаков, уникальных для Nothobranchius и не встречающихся у Paranothobranchius. Так как они не могли доказать монофилетичность Nothobranchius, они предпочли не считать Paranothobranchius самостоятельным родом, так как это продразумевало бы родство сестринских групп. Сестринская група Paranothobranchius может быть так же другой линией внутри рода Nothobranchius.
Типовой материал
В своём переописании N.microlepis, Wildekamp & Haas (1992) образовали лектотип для этого вида. Описание базировалось на синтипичных экземплярах N.microlepis и на более поздних коллекциях из Сомали.
Vinciguerra (1897) утверждал, что все анализированные типовые экземпляры были сильно деформированы. Внутренности, жаберные дуги и жаберные мембраны у некоторых были удалены. Это не давало возможности произвеси правильные измерения, что может объяснять расхождение данных, полученных Wildekamp & Haas и другими (Wildekamp & Haas, 1992).
Лектотип
Museo Civico di Storia Naturale, Genoa [MCSNG] 15163: 1 самец 52.4 mm SL, найденный V. Bottego 31 X 1895 в воде в застойной воде у подножия горы Egherta, между Brava и Lugh [ныне Lugh Galana] [около 2° 20' N - 43° 20' E] by.
Паралектотипы
- MCSNG 15163: 4 самца и 3 самки 49.3-61.8 mm SL, найдены там же, где и лектотип.
- MCSNG 15163: 12 экземпляров, выловлены там же где и лектотип.
Дополнительный материал
Дополнительный материал, исследованный Wildekamp & Haas (1992) составляет:
- MCSNG 47886: 6 самцов и 6 самок 39.0-50.0 mm SL, Warfa [02° 54'N - 44° 13' E], Somalia, выловлены Haas 30-I-1979.
- BMNH 1983.3.14: 74-75, 1 самец и 1 самка 45.7-47.3 mm SL, Durbane [02° 59' N - 44° 19' E], Somalia, выловлены I. Aweis и A. Ibrahim 24-V-1982.
- BMNH 1983.3.14: 76, 1 самец 55.0 mm SL, Warfa [02° 54'N - 44° 13' E], Somalia, выловлен WHO, 1983.
- BMNH 1987.1.19: 1-7, 4 смца и 3 самки 33.3 - 44.9 mm SL, около Warfa [02° 54'N - 44° 13' E], Somalia, выловлены R. Wildekamp & A. Ibrahim 6-VI-1983.
- MRAC 84-50-P-35-45, 6 самцов и 5 самок 32.5-45.5 mm SL, около Warfa [02° 54'N - 44° 13' E], Somalia, выловлены R. Wildekamp & A. Ibrahim 6-VI-1983.
- MRAC 86-48-P-1-10, 5 самцов и 5 самок 37.3-44.0 mm SL, Bur Eibi [02° 59' N - 44° 18' E], Somalia, выловлены R. Haas 30-I-1979.
- MRAC 86-48-P-26-32, 5 самцов и 2 самки 25.0-31.5 mm SL, Hogay [02° 54' N - 44° 14' E], Somalia, выловлены R. Wildekamp & A. Ibrahim 6-VI-1983.
Морфометрические и меристические данные
D= 16-17; A= 17-18; количество чешуй в боковом продольном ряду: 40-42.
(чешуйки в боковом ряду сложно подсчитать, так как они мелкие и расположены неупорядоченно).
Кариотип: кариотип N.microlepis из Warfa состоит из 12 хромосом, все хромосомы набора телоцентрические, кроме одного крупного метацентрического элемента. n = 12, 2n = 24, 13 плеч, сравнительная длина самого длинного плеча равна 17,4% от суммы всех плеч (J. J. Scheel, pers. comm. to Wildekamp & Haas, 1992).
Wildekamp & Haas (1992) рассмотрели это определение вида: его можно отличить "от всех остальных видов рода по горбу, изогнутой линии спины и резкому переходу между спиной и головой, большому количеству мелких и неупорядоченно расположенных чешуек в продольных рядах, 26 - 30 жаберных тычинок против 12 - 15 у остальных видов, длинных и близко расположенных, тёмной вертиуальной полосе, идущей через глаз, однотонной тусколой окраске самца и поведению во время ухаживания".
| место лова | Egherta | Bur Eibi | Warfa | Hogay | Durbane | ||||||||
| разброс | X | SD | разброс | X | SD | разброс | X | SD | разброс | X | SD | разброс | |
| SL | 49.3-52.4 | 38.8-44.0 | 35.3-55.0 | 25.0-31.5 | 47.3 | ||||||||
| BD | 31.7-35.7 | 33.7 | 1.32 | 28.1-30.2 | 29.3 | 0.77 | 27.2-33.7 | 30.7 | 1.83 | 23.6-27.4 | 25.7 | 1.48 | 28.5 |
| HL | 29.0-33.3 | 30.6 | 1.57 | 31.2-34.2 | 32.6 | 1.24 | 30.0-33.7 | 31.2 | 1.22 | 30.0-33.2 | 31.7 | 1.03 | 33.0 |
| ED | 6.2-7.1 | 6.7 | 0.39 | 7.5-8.2 | 8.0 | 0.26 | 6.4-8.4 | 7.2 | 0.50 | 7.4- 7.7 | 7.6 | 0.10 | 6.8 |
| IW | 13.9-15.2 | 14.6 | 0.50 | 11.8-13.6 | 12.5 | 0.67 | 11.7-15.4 | 13.1 | 0.79 | 11.2-12.7 | 12.1 | 0.64 | 12.9 |
| SnL | 6.7-7.7 | 7.3 | 0.34 | 7.6-9.3 | 8.1 | 0.65 | 6.5-8.5 | 7.6 | 0.56 | 6.8-9.7 | 7.9 | 1.03 | 7.2 |
| PDL | 54.8-59.3 | 56.0 | 1.48 | 57.0-61.3 | 58.5 | 1.55 | 55.8-63.5 | 58.4 | 1.85 | 56.2-61.0 | 58.9 | 1.75 | 55.0 |
| PAL | 59.3-61.5 | 60.4 | 1.28 | 53.3-57.0 | 55.1 | 1.40 | 52.0-62.3 | 55.7 | 2.53 | 56.2-58.7 | 57.2 | 0.83 | 52.9 |
| PPL | 45.4-47.8 | 46.9 | 0.96 | 41.2-45.9 | 44.0 | 1.92 | 42.0-50.2 | 45.3 | 2.09 | 45.7-49.2 | 47.9 | 1.22 | 42.1 |
| CL | 18.9-19.8 | 19.3 | 0.35 | 18.0-20.4 | 19.1 | 0.87 | l8.2-21.7 | 19.8 | 1.01 | 17.7-20.1 | 18.8 | 1.05 | 17.8 |
| CD | 12.0-15.3 | 13.7 | 1.52 | 12.9-13.8 | 13.4 | 0.37 | 12.7-15.9 | 14.2 | 0.99 | 11.8-13.3 | 12.7 | 0.57 | 13.7 |
| n | 5 | 5 | 17 | 5 | 1 | ||||||||
самцы
| место лова | Egherta | Bur Eibi | Warfa | Hogay | Durbane | ||||||||
| разброс | X | SD | разброс | X | SD | разброс | X | SD | разброс | X | SD | разброс | |
| SL | 41.7-48.3 | 37.3-40.4 | 32.5-43.0 | 25.0-27.2 | 45.7 | ||||||||
| BD | 29.8-30.6 | 30.3 | 0.36 | 26.3-29.0 | 27.5 | 0.92 | 26.0-32.6 | 29.6 | 2.06 | 22.0-22.8 | 22.4 | 0.40 | 25.8 |
| HL | 29.8-32.5 | 31.0 | 1.12 | 32.2-35.0 | 33.6 | 0.97 | 28.0-33.9 | 30.7 | 1.54 | 30.0-31.3 | 30.7 | 0.65 | 29.3 |
| ED | 6.0- 7.0 | 6.6 | 0.45 | 8.3-9.4 | 8.9 | 0.37 | 6.8-8.6 | 7.9 | 0.55 | 8.0-8.1 | 8.1 | 0.05 | 7.0 |
| IW | 12.5-14.5 | 13.4 | 0.84 | 11.1-12.4 | 11.8 | 0.48 | 9.4-13.2 | 11.6 | 1.10 | 12.0-12.1 | 12.1 | 0.05 | 10.9 |
| SnL | 7.9-9.1 | 8.5 | 0.49 | 7.5-8.1 | 7.8 | 0.21 | 6.3-8.4 | 7.1 | 0.65 | 6.8-7.3 | 7.1 | 0.25 | 6.6 |
| PDL | 57.0-63.1 | 59.8 | 2.52 | 59.4-63.4 | 60.0 | 3.08 | 59.5-66.1 | 62.2 | 1.83 | 60.0-62.5 | 61.3 | 1.25 | 61.3 |
| PAL | 62.0-64.4 | 63.6 | 1.11 | 55.7-65.2 | 62.8 | 3.55 | 58.8-66.8 | 63.5 | 2.00 | 60.0-64.3 | 62.3 | 2.15 | 59.1 |
| PPL | 46.4-50.8 | 48.7 | 1.80 | 44.6-49.2 | 47.6 | 1.62 | 42.6-52.6 | 49.5 | 2.49 | 48.0-49.3 | 48.7 | 0.65 | 46.6 |
| CL | 16.3-18.5 | 18.0 | 0.94 | 17.6-22.0 | 19.4 | 1.47 | 17.5-20.9 | 19.1 | 0.92 | 18.4 | 18.8 | ||
| CD | 13.5-14.3 | 14.0 | 0.33 | 11.7-13.4 | 12.5 | 0.63 | 11.6-14.1 | 13.1 | 0.76 | 12.5 | 13.6 | ||
| n | 3 | 5 | 14 | 2 | 1 | ||||||||
самки
Таб 1: Морфометрические характеристики N.microlepis (выражены в % от SL) (по Wildekamp & Haas, 1992).
[SL: стандартная длина; BD: высота тела ; HL: длина головы; ED: диаметр глаза;
IW: межглазничное расстояние; Snl: длина рыла; PDL: предорсальная длина;
PAL: преанальная длина; PPL: превентральная длина; CL: длина хвостового стебля;
CD: высота хвостового стебля; n: количество измеренных экземпляров]
| вид | место лова | спинной плавник | анальный плавник | ||||||||||||
| 16 | 17 | 18 | 19 | 20 | X | 16 | 17 | 18 | 19 | 20 | 21 | 22 | X> | ||
| N.microlepis | Egherta | 4 | 3 | 16.6 | 2 | 5 | 1 | 17.9 | |||||||
| Bur Eibi | 2 | 3 | 2 | 1 | 17.7 | 4 | 4 | 2 | 18.8 | ||||||
| Warfa | 9 | 18 | 4 | 17.3 | 9 | 20 | 16 | 1 | 18.2 | ||||||
| Hogay | 1 | 4 | 17.1 | 1 | 5 | 1 | 18.0 | ||||||||
| Durbane | 2 | 17.0 | 2 | 18.0 | |||||||||||
| N.fasciatus | Goba | 4 | 2 | 17.3 | 4 | 2 | 19.3 | ||||||||
| Haya | 6 | 6 | 2 | 16.7 | 1 | 2 | 5 | 4 | 2 | 18.3 | |||||
| Afmadu | 1 | 18.0 | 1 | 20.0 | |||||||||||
| N.bojiensis | 8 | 6 | 1 | 18.5 | 8 | 2 | 4 | 1 | 19.9 | ||||||
| вид | место лова | количество чешуй | |||||||||||||
| 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | 40 | 41 | 42 | 43 | 44 | X | ||
| N.microlepis | Egherta | ||||||||||||||
| Bur Eibi | 1 | 2 | 5 | 40.8 | |||||||||||
| Warfa | 2 | 2 | 4 | 1 | 1 | 37.5 | |||||||||
| Hogay | 6 | 7 | 6 | 10 | 9 | 4 | 4 | 38.8 | |||||||
| Durbane | 2 | 1 | 1 | 3 | 40.7 | ||||||||||
| N.fasciatus | Goba | 1 | 1 | 1 | 2 | 1 | 37.5 | ||||||||
| Haya | 1 | 4 | 3 | 2 | 3 | 1 | 35.4 | ||||||||
| Afmadu | 1 | 38.0 | |||||||||||||
| N.bojiensis | 1 | 3 | 3 | 1 | 3 | 4 | 41.9 | ||||||||
Таб.2 :Количество спинных и анальных плавников и чешуй в продольных рядах у N.microlepis, N.fasciatus и N.bojiensis (по Wildekamp & Haas, 1992)
Описание
N.microlepis явялется замечательным представителем рода. Он отличается от остального большинства Nothobranchius формой тела - наличием горба у взрослых самцов, большим количеством мелких чешуй, неяркой окраской и характериной вертикальной тёмной полосой, идущей через глаз, начиныющаяся выше глаза и доходящая доуголка рта (Haas, 1982). Хвостовой плавник у самцов имеет тонкую чёрную кайму, самца значительно равьируют по форме тела - у некоторых сильно выраженный горб перед глазом, а у других почт нет его. (Haas, 1982). Анальный плавник у самок сильно удлиннён, от тела до кончика вдвое больше чем у других Nothobranchius и заметно длиннее чем у самцов. (Haas, 1982). Так же от остальных Nothobranchius этот вид отличается поведением. Во время ухаживания самцы стараются коснуться своим "горбом" брюха самки. Самец приближается к самке снизу в отличие от того, что наблюдается у других видов Nothobranchius. Самки сильно отличаются от других видов удлинённым первыми анальными лучами, первые из которых жёсткие и ветвятся только около кончиков (Wildekamp, 1984). Это способствует более глубокому погружению икры в грунт. Жёсткий и неветвящийся первый анальный луч так же имеется у самок N.virgatus, населяющего сходные болотистые места.
Wildekamp & Haas (1992) провели переописание этого вида. Чешуя на теле мелкая и неупорядоченно расположенная, особенно в передней части тела. Все чешуйки в среднем продольном ряду имеют неглубокие ямки, не связанные с леащими глубже органами боковой линии. Передняя часть головы, рыло, губи и подбородок голые. Предкрышки, жаберные крышки и щёки покрыты очень мелкими чешуйками. Рыло умеренно удлинённое, тор субтерминальный, слегка выступающий и широко раскрывающийся. Врхняя челюсть выступает на уровне переднего края зрачка. На верхней и нижней челюстях имеется большое количество онких конических зубов, погружённых в губчатую ткань. Зубы внешнего ряда более крупные, более выступающие на верхней челюсти. Жаберная мембрана выступает за жаберную крышку, свободный край бахромчатый. На первой жаберной дуге 26 - 30 жаберных тычинок, длинных и густо расположенных. [Figure 4]. Передняя и центральная система органов боковой линии расположены в двух мелких канавках, по шесть органов в каждой. Задние системы отрганов боковой линии сильно изогнуты под углом менее 90о с четыремя органами в каждой. Преоперкулярная система рганов боковой линии располагается в отдельной открытой канавке, её задний край заходит за жаберную крышку.
Самцы имеют крепкое телосложение и, по сравнению с другими Nothobranchius, тело сжато с боков и более крупные, достигают 60 мм SL. Линия спины сильно искривлена, наклон сильно меняется над глазом [Figure 5]. Большинство взрослых самцов имеют горб и наибольшая высота тела составляет 3,0 - 3,8 единиц от стандартной длины за жаберной крышкой. У более молодых самцов горб отсутствует или менее выражен. Имеется узкий спинной гребень между задней частью головы и началом спинного плавника. Голова очень большая, 2,9 - 3,3 единиц от SL. Спинной и анальный плавники округлые, у более взрослых самцов кончики лучей выступают за мембрану. Проксимальная часть покрыта эпидермальной тканью. На последних 5 - 6 спинных лучах имеются мелкие папиллы. Папиллы на всех анальных лучах более крупные. Хвостовой плавник треугольный. У взрослых самцов только в основании грудных плавников, нижней части тела и жаберных крышках на чешуйках имеются ктеноидные шипики. Чешуя на задней части тела как правило покрыта эпидермальной тканью, ктеноидные шипики выдающиеся (Wildekamp & Haas, 1992).
Самки мельче самцов того же возраста, достигают 50 мм SL, высота тела меньше, 3,1 - 4,5 единицы от SL. Линия спины слегка искривлена, наклон начинается над или непосредственно перед глазом. Нибольшая высота тела находится над брюшными плавниками. Голова так же крупная - 2,9 - 3,6 единиц от SL. Спинной плавник округлый, квостовой треугольный, анальный длинный и заострённый с длинными и жёсткими 3 - 12-ю передними лучами, ветвящимися только у кончика. Ни стеноидных шипиков, ни папилл на плавниках не имеется. Эпидермальной ткани на плавниках или чешуйках задней части тела нет. Жаберная мембранаслегка выдаётся за край крышки, свободный край не бахромчатый (Wildekamp & Haas, 1992).
Окраска
По (Wildekamp & Haas, 1992).
У самцов тело однотонно коричневато-голубое, довольно блёклое. Свободные края чешуек с узкой кричневой каймой. В основном, у большинства особей нет рисунка, но у некоторых может быть 3 - 4 короткие тёмно-серые очень узкие полосы между грудными и брюшными плавниками. Брюхо светло-серое. Спинной плавник серо-голубой, более тёмный ближе к краю. В основании имеется мраморный рмунок из серо-коричневых крапинок, становящихся мельче ближе к краю, параллельных плавниковым лучам. Анальный - серо-голубой, без рисунка, но в основании более светлый. Может присутствовать светло-коричневый или жёлтый оттенок. Брюшные плавники серо-голубые, на концах становящиеся очень тёмными, почти чёрными. Хвостовой плавник серо-голубой, с небольшими коричневыми точками на верхней части; ближе к основанию обычно имеется заметная узкая полоска по краю. Заметная тёмно-серая или чёрная полоса идёт от верхней части головы через глаз к углу рта. Выступающая часть жаберной мембраны розовая, на бахромчатом крае имеется узкая кремовая кайма. (Wildekamp & Haas, 1992).
Самка: Тело однотонно коричневато-серое, иногда с бледным зелёно-голубым оттенком, довольно светлым. Центральная часть чешуек передней части тела и спине более светлого коричнево-серые до желтоватых. Брюхо светло-серое. На теле нет рисунка и отсутствует тёмная полоса, идущая через глаз. Спинной и хвостовой плавники абсолютно прозрачные и бесцветные. Анальный прозрачный, жёлто-коричневый у основания, блёкло-белый около кончика (Wildekamp & Haas, 1992).
Размер
В аквариуме самца достигают максимальной длины 75 мм; самки остаются на 20 - 25 мм мельче.
Код
MIC
Распространение и биотоп
Wildekamp & Haas (1992) сообщают, что V. Bottego во время второй его экспедиции в Сомали и Gallaland вылавливал рыб у подножия Egherta, большой горы в кутарниковой саванне. Пруды, в которых был найден N.microlepis содержали дождевую воду и пересыхали через два месяца после сезона дождей. Это место состетствует прежней деревне Egherta, находящейся в дренажной системе пересыхающей реки Madagoi [так же Bohol Matagoi] в районе Bay в южной Сомали (Wildekamp & Haas, 1992).
Весь ареал N.microlepis пока ещё не полностью определйн, но недавнние коллекции в природе в южной Сомали и последующие в северо-всточной Кении показали, что он шире, чем предполагался ранне. Ареал включает большое количество временных водоёмов вокруг верхнего и среднего течения системы реки Uebi Scebeli River. Может сложиться неправильное впечатление, что эта река является единственной, где во временных водоёмах процветает этот вид. Всё же, все известные водоёмы питаемы дождевой водой. Недаввно было идентифицировано место поимки этого вида в системе реки Tana в Кении. Известно ещё одно недавнее место вылова в южной Кении [дорога Mariakani-Samburu].
Haas (1981) в январе - феврале 1979г. выловил N.microlepis закрыл неописанный вид N. spec. "Warfa-Blue" [впервые описанны как N.cyaneus, в настоящее время считающийся более поздним синонимом N.jubbi] в большом углублении ["вар" = "Whar"], заполненом дождевой водой. Это место находилось примерно в 180 км на северо-запад от Mogadishu, около деревни Warfa в кустах, примерно в 18 км восточнее большого массива Bur Hacaba [или Acaba] на дороге из Mogadishu в город Baidoa. В этой местности, на боее чем 150 км удалённой от ближайшей реки или какого-либо другого постоянного водоёма, дождевая воды собирает в больших углублениях, увеличенных местными жителями для сбора воды. Тавие "вары" ("War") по сравнению с придорожными канавами находящимися южнее и где Haas (1981) находил N.patrizii, были полностью лишены растительности. Вода в "варах" была аболютно непрозрачной с красноватым оттелком. Дно состояло из красной глины, температура воды составляла от 26.5 до 31°C [80-88 °F], наивысшая - около берегов; рН - около 7,8, dGH 36-39. Haas (1981) был информарован о том, что большинство "варов", за исключением очень сухих лет, более или менее пересыхали на более чем 2 месяца, в центре располагалось большое углубление, где вода оставалась доступной в очень сухое время. В январе 1979 водоём в Warfa был примерно 300 х 100 м и 2,5 м глубиной.
Dr. Haas подсчиатл, что такие "вары" за исключением черезвычайно засушливых лет, не должны пересыхать более чем на 2-3 месяца. Haas (1981) не привёз живых рыб найденного им вида, но вместо этого сообщил, что N.patrizii, N.microlepis и N.cyaneus "Warfa-Blue" были привезены в США в 1979г., так как икра была собрана вместе с илом в их временных водоёмах.
Haas (1981) уделил внимание в середине 1982 на тот факт, что Сомали - очень сложная для въезда и передвижения внутри неё страна и что таким образом шансы на то, что кто-то ещё, интересующийся мелкими рыбами будет иметь возможность посетить эту страну, очень малы. Haas рекомендовал прилагать максимум усилий для распространения этих рыб среди любителей, так как эти три вида привезённые в икре в иле в 1979 г. [N.microlepis, N.patrizii и N.cyaneus / jubbi "Warfa-blue"] следует считать редкими в хобби. Всё же, в начале 90-х годов N.microlepis полностью исчез из хобби, N.cyaneus/jubbi "Warfa-blue" стал очень редким у любителей, и только N.patrizii очень широко распространился. Почему не увенчались успехом попытки хорошо организованной Ассоцииции Любителей? N.microlepis выживал в хобби всего несколько лет.
Wildekamp посетил местность между Bur Acaba и Bur Eibi, вкючая Warfa, 6 июня 1983 и обнаружил N.microlepis в пяти "варах" [Dekta, Warfa, Hogay, Kaisany и Durbane]. Все эти водоёмы были заполнены дождевой водой. Только "вар" в Hogay был почти полностью высохшим (оставалось около 10-cm воды). Во время визита Wildekamp в Warfa "вар" был больше, около 400x 150 м шириной, с температурой 33.5°C [92°F], DGH 8°, DKH 7° и pH 6.7. Wildekamp & Haas (1992) объясняют различия в жёсткости и рН между I-1979 и VI-1983 большим периодом испарения и соответственно концентрации минеральных веществ. Так же во время последнего визита ни один из "Варов" на содержал водной растительности, только частично берега бали поросши травой, частично заходя в воду. За исключением вара в Warfa, где дно состоялло из серого ила и латерита, во всех остальных дно было илистое с большим содержанием гумуса.
Wildekamp (1983(a) приводит короткое замечание о встречаемости видов Nothobranchius в соответствии с основным составом почв. Forstner и Willert (Willert, 1984) обнаружили одинаковый состав почвы в местах обитания N.microlepis вдоль дороги из Garsen в Garissa в северной Кении. 30 июня 1983г. пруд в 53-х км от Garsen был около 20 см глубиной и 8х4 м шириной был заполнен мутной серой водой. Его почва состаяла из серого ила, водной раститенльости не было. Сходная ситуация обнаружилась и в другом пруду, в 1 км дальше по дороге (M. Forstner, pers. comm. to Wildekamp & Haas, (1992)). S. Baer (pers. comm. to Wildekamp & Haas, (1992)) сообщил им о временном пруде, 3.5-км на юге по главной дороге из Mombasa в Nairobi, примерно посередине между Mariakani и Samburu. Во всех местах обитаниия, за исключением последнего, в которых встречался пока ещё неопределённый вид Nothobranchius, N.microlepis обитает синтопично с голубой формой N.jubbi Wildekamp & Berkenkamp, 1979 [ранее описанный как N.cyaneus Seegers, 1981]. Двоякодышаие рыбы Protopterus amphibius (Peters, 1844) был обнаружен синтопично в Warfa и Hogay.
В июне - июле 1983г. M. Forstner, Manfred и Brigitte Willert предприняливоезку в Кению. Их база располагалась в Mombasa. Оттуда проводились азличные выезды для ловли рыб. Во время одной из таких поездок на север в сторону дренажной сисетмы реки Tana River они обнаружили в 53 км севернее Garsen, в направлении Garissa, N.cyaneus вместе с N.microlepis.
Между концом мая и серединой июля 1983г. Wildekamp (1984) провёл миссию для World Health Organisation [WHO] в северной Сомали. Между деревнями и монолитами Bur Acaba и Bur Eibi, он выловил в нескольких "варах" [природных углублениях, заполняемых дождевой водой во время сезона дождей или прудах, вырытых людьми для запасания дождевой воды]. Wildekamp (1984) проверил несколько "варов" около деревень Dekta [точка S-1], Warfa [S-2], Hogay [S-3], Kaisany [S-4] и Durbane [S-5]. Все эти вары содержали одинаковую фауну Nothobranchius, а именно N.microlepis и N.cyaneus, для которого N.spec. "Warfa blue" является синонимом.
Wildekamp (1984) заметил, что N.microlepis всегда встречается в больших количествах. Они явно предпочитают держаться в глубоких местах, хотя это и не является правилом. Вода в этих изолированных водоёмах всегда была взмученной, варьируя от сероватой до красновато-коричневой, в зависимости от состава местной почвы. Общая жёсткость варьировала от 5 до 15 DH, карбонатная - от 4 до 6 DH и рН около нейтральной 6,3 - 8,3. При измеренных высоких температурах воды (всегда около 30° C - 86° F), количество растворённого кслорода была еслественно довольно низкой между 3 и 5,5 мг/л. Самая высокая жёсткость и рН были отмечены в водоёме S-3, около Hogay, в "варе", который во время псещения почти полностью пересох. В этом водоёме , количество рыб было большим, но все они были в доольно хорошем состоянии. В качестве сопутствующих видов Wildekamp (1984) обнаружил почти везе Protopterus amphibius (Peeters, 1844) [местное название Ramole], лягушек, водных насекомых, большое количество пресноводных крабов и водяных скорпионов. Несмотря на присутствие нескольких хищников, Wildekamp (1984) отметил, что плотность рыб была везде довольно высокой, по некотрым оценкам она достигала около 20 экземпляров/л.
В южной Сомали, ближе к границе с Кенией и несколько км от деревни Goba Wildekamp (1984) обнаружил ещё один вид Nothobranchius, похожий на N.microlepis (большое количество мелкой чешуи, жёсткий первый анальный луч у самок, чёрная полоса через глаз). Догога была опять затоплена и глубина воды составляла примерно 1м. Машина не могла проехать вокруг, так как с одной стороны находилось болото, а сдругой - густые колючие кусты. Пока машина стояла из-за закипания двигателя Wildekamp (1984) начал ловить со своего сиденья и выловил самку. У неё были вертикальные чёрные штрихи на боках, но не похожие на самок некоторых популяций N.melanospilus (Pfeffer, 1896). Через некоторое время был пойман и соответствующий самец. Этот вид, предварительно названный "N.spec. affinis microlepis Goba B", теперь известен как N.fasciatus Wildekamp & Haas, 1992. Цвет тела серо-голубой, непарные плавники более тёмные серо-голубые. На задней части тела имеются поперечные тёмные серые штрихи. Чёрная маргинальная полоса на хвостовом плавнике, имеющаяся у N.microlepis, у N.fasciatus отсутствует.
Судя по музейным экземплярам, большинство рыб было выловлено с отября по июнь. В 1895 лектотип и паралектотипы были выловлены 31 октября; Haas выловил взрослых особей 30-I-1979; Aweis & Ibrahim поймали рыб 24-V-1982; и Wildekamp & Ibrahim - 6-VI-1983.
Ott (1992) приводит характеристики, встречающиеся в природных водоёмах: жёсткость между 36 и 39° dGH; pH - 7.8; температура воды 27 - 31° C. в январе и dGH 8°; 33.5° C и pH 6.7 в июне.
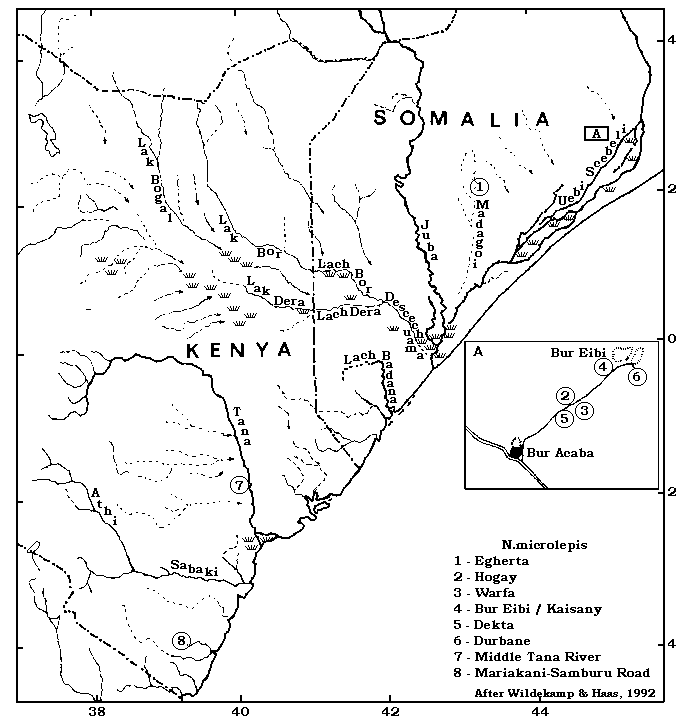
Рис 1: Распространение N.microlepis в Сомали и Кении (по Wildekamp & Haas (1992).
Содержание и разведение
Примерно век N.microlepis оставался неизвестен для любителей. Поэтому, тот факт, что во время научной работы в Сомали Dr. R. Haas в 1979г. обнаружил популяцию этого вида вызвало большой интерес. Открытие дало ему возможность исследовать и описать место обитания дикого N.microlepis и отловить несколько особей и даже икру этого редкого вида. (Jubb, 1981).
Всё же N.microlepis возсожно один из наименее привлекательных видов рода. Даже опытные любители, разводящие Nothobranchius, считают этот вид сложным в содержании и разведении. Эти рыбы никогда не были широко распространены у любителей, и следовательно исчез вскоре после нескольких привозов.
По данным Haas (1981), никаких необычных требований по сравнению с другими родственными видами эти рыбы не требуют. Они нерестятся в торф, инкубация занимает 8 - 12 недель при температуре около 24°C [74°F]. Молодь растёт быстро. Первые признаки пола проявляются в возрасте около 18 дней, а первые спаривания могут начаться в 24 - 26 дней. N.microlepis достигает 3,5 дюйма самцы и 3,25 дюймов самки.
Но, по данным Wildekamp (1983b), разведение часто неуспешно и через 1 неделю икрметания большая часть икры подвергается атаке грибков и погибают из-за нарушений в процессе деления клеток. Причины этого неизвестны. Здоровая икра развивается 3 - 4 месяца. Малёк во время выклева сравнительно крупный и сразу может поедать только что выклюшувшихся науплиусов Артемии. Они нуждаются в больших количествах корма, так как его нехватка приводит к необратимым нарушениям роста. При достаточном количестве кормов они могут достигнуть половой зрелости к 6 неделям.
В аквариумах, принадлежащих Ministry of Health в Mogadishu, Wildekamp (1984) несколько раз наблюдал, что самки N.microlepis может съесть более 350 личинок комаров в день. Исследования содержания желудков выловленных в природе показало однако, что личинки комаров не являются главным компонентом рациона N.microlepis. Он состоит в основном из большого количества науплиусов мелких ракообразных. Так как строение, расположение и размер рта, так же как и размер и строение зубов, показывают, что добыча может быть и более крупной, что так же было проверено. Как можно было ожидать, Wildekamp (1984) отметил, что более крупные личинки насекомых и даже мелкие особи N.cyaneus из того же биотопа так же могут служить добычей. Следует всё же заметить, что N.microlepis можно считать очень прожорливым видом. Случайное поедание мелких раб происходило при случайном кормлении провёрнутым козьим сердцем. (Wildekamp, 1984).
Обычно при нересте Nothobranchius самец находится немного сверху. Затем самец направляет самку ко дну, где происходит спаривание, когда он прижимается к ней своим горлом в райне передней части головы (Haas, 1976). Биология размножения N.microlepis заметно отличается от этого (Foersch, 1984). Самец приближается к самке снизу, равком прижимается горбом к брюшку самки. Это приглашает её в сопровождении самца плыть вниз ко дну, где происходит обычное для Nothobranchius икрометание. Самец обнимает самку спинным и анальным плавниками, самка погружает свёрнутый в трубочку анальный плавник в торф. После откладывания икринки, партнёры расходятся в координированном движении, названном Haas (1976) "толчком". Это движение заканчивается тем, что икринка поклывается взмуженным субстратом. Икрометание повторяется много раз, обычно чаще, чем у других видов рода и заканчивается бегством самки. Затем самец может продолжить поиски следующей готовой самки. Обычно икрометание происходит в середине дня (Wildekamp & Haas, 1992).
Wildekamp & Haas (1992) наблюдали, что приближение и контакт самца снизу является заметным отклонением от обымного контакта у всех известных видов Nothobranchius и предположили значительную сенсорну функцию увеличенного горба самца. Всё же сходное "приближение снизу" наблюдается у самцов N.flammicomantis, у которого нет развитого горба на спине.
Foersch (1984) отмечает, что мальки в момент выклева очень крупные и сразу могут поедать науплиусов Артемии, через 4 дня гриндальского червя, через 11 - мелкого трубочника и через 16, при длине 3 см, мелкого матыля! Несколькими днями позже можно распознать пол рыб (по более длинному анальному плавнику у самок), после чего рыб разных полов рекомендуется рассадить. Через 5 недель, мальки достигают примерно 5,8 см длины и вырастают до 6,8 - 7 см ещё через неделю. В возрасте 4 - 5 недель самцы начинают окрашиваться. В это же время начинает проявляться тонкая чёрная кайма на дистальной части хвостового плавника. Большой горб на уровне горла перед глазом начинает развиваться в следующие недели, но не у всех особей - самцов (Foersch, 1984). Взорслых рыб так же следует обильно кормить, в противном случае их состояние здоровья быстро ухудшается.
В связи со значительной активностью самцов и бурным процессом спаривания лучше содержать рыб разных полов раздельно и сажать вместе для нереста на короткий период, а не содержать их постоянно в нерестовике. (Foersch, 1984) сажал рыб вместе после через 2-3 дня и за один раз получил 191 икринку после такого короткого раздельного содержания. Самки не так плоловиты как у других однолетних рыб и период икрометания короче чем у других видов Nothobranchius. У большого количества икру наблюдаются нарушения развития. (Foersch, 1984) контролировал развитие около 400 икринок: через 3,5 месяца выклюнулось 55 мальков; 47 икринок имели довольно продвинутое развитие эмбриона, 2 оставались прозрачными без признаков развития. Ещё черех месяц инкубации выклюнулись дополнительные 15 мальков. Все остальные икринки погибли и разложились.
Wildekamp (1983b) наблюдал особенное поведение группы почти взрослых рыб. Они часто плавали друг за другом над дном поисках корма.
Этот вид рекомендуется для опытных любителей и его не следует заводить новичкам.
Литература
 : 122-125, 5 figs.
: 122-125, 5 figs.
© Авторское право 2000, Marc Bellemans. Все права зарезервированы. Оригинал статьи
- Войдите на сайт для отправки комментариев
Новые обсуждения форума
- Группы в месенджерах
- Выставки Killi в 2025 году (РЕЗУЛЬТАТЫ)
- Предновогодняя встреча 2024.
- Выставки Killi в 2024 году (РЕЗУЛЬТАТЫ)
- Февральская встреча 9 числа
- Поздравляем!
- Астролебиасы (Austrolebias)
- Лягушатники - тоже люди?
- Выставки Killi в 2023 году (РЕЗУЛЬТАТЫ)
- Sergejam Dzim?anas Diena!
- "Моно" аквариум самосевом.
Активные обсуждения форума
- Продам эпиплатис Дагета .
- Группы в месенджерах
- Выставки Killi в 2025 году (РЕЗУЛЬТАТЫ)
- Предновогодняя встреча 2024.
- "Моно" аквариум самосевом.
- Выставки Killi в 2024 году (РЕЗУЛЬТАТЫ)
- Общая выставка с нашим участием.
- Февральская встреча 9 числа
- Сведения о биотопах
- Александр Сухов с днём рождения!
- Поздравляем!
Новые записи в блогах
- Всем привет. Немного психотерапии для нашего сообщества
- Epiplatys dageti
- Один из способов фотографирования килли
- Fundulopanchax walkeri spurelli
- N. Rachovii
- Aphyosemion australe фото дня
- Fundulopanchax gardneri "Gold" фото дня
- Аквариумные Аплохейлусы
- Разведение Fundulopanchax gardneri nigerianus
- Про выставку аквариумных рыб